
{kind=link}
抗菌肽SAMP1及其类似肽的构效关系
[常俊朋, 赵佳瑞, 陈思佳, 孟凯, 石微妮, 李瑞芳 ]
]
]
|
|


联系人简介: 李瑞芳, 女, 博士, 教授, 博士生导师, 主要从事多肽药物方面的研究. E-mail: lrf@haut.edu.cn
以人工合成抗菌肽1(Synthetic antimicrobial peptide 1, SAMP1)为研究模板, 采用氨基酸序列重排、 不同的带正电荷氨基酸残基和疏水性氨基酸残基取代等方法, 设计合成了8条SAMP1类似肽. 利用生物信息学软件预测了SAMP1及其类似肽的理化性质; 采用圆二色光谱(CD)技术测定其在不同环境下二级结构的变化; 采用噻唑蓝(MTT)法测定其抗菌活性; 通过红细胞溶血实验评估了这些多肽的溶血性. 结果表明, 大部分类似肽具有较低的溶血毒性和较高的广谱抗菌活性. CD光谱分析结果显示, 大部分类似肽二级结构以α螺旋和无规则卷曲为主, 在体积分数为50%的2,2,2-三氟乙醇(TFE)溶液中, α螺旋结构比例增加. 与母肽SAMP1相比, 经序列重排后得到的SAMP1-A1, SAMP1-A2和SAMP1-A3的抗菌活性变化不大, 但序列中正电荷氨基酸残基均匀分布的类似肽SAMP1-A2的溶血毒性增加. 用精氨酸(Arg)取代SAMP1序列中的赖氨酸(Lys)得到的类似肽SAMP1-A4的抗菌活性增强, 同时溶血毒性降低. 用疏水性较强的异亮氨酸(Ile)和缬氨酸(Val)取代SAMP1中的疏水性氨基酸残基, 得到的类似肽SAMP1-A5和SAMP1-A7的抗菌活性急剧降低; 用疏水性较弱的色氨酸(Trp)取代SAMP1中的疏水性氨基酸残基, 得到的类似肽SAMP1-A8的抗菌活性增强, 同时溶血毒性提高.
The synthetic antimicrobial peptide 1(SAMP1) was used as the research template, eight SAMP1 analog peptides were designed and synthesized by amino acid sequence rearrangement, different positive charged amino acids replacement and hydrophobic amino acid substitutions. Bioinformatics software was used to predict the physicochemical properties of SAMP1 and its analog peptides. Secondary structures were determined by circular dichroism(CD) technique. Antimicrobial activity was determined by 3-(4,5-dimethylthiazolyl)-2,5-diphenyltetrazoliumbromide(MTT) assay. Hemolytic properties of peptides were evaluated by erythrocyte hemolysis assay. The results showed that most of the analogues of SAMP1 had low hemolytic toxicity, and high broad-spectrum antimicrobial activity. The results of CD showed that their secondary structures were mainly α-helix and random coil, the proportion of α-helix increased in the 50% volume fraction of 2,2,2-trifluoroethanol(TFE) solution. Compared with mother peptide SAMP1, the antimicrobial activities of the analog peptides SAMP1-A1, SAMP1-A2 and SAMP1-A3, which were obtained after sequence rearrangement, did not change so much. While the hemolytic toxicity of SAMP1-A2, in which uniform distribution of positive charged amino acids in the sequence, increased. The antimicrobial activity of the peptide SAMP1-A4, which was obtained by substituting arginine(Arg) for lysine(Lys) in the SAMP1 sequence, improved with less hemolytic toxicity. The antimicrobial activity of the peptides SAMP1-A5 and SAMP1-A7, obtained by replacing the hydrophobic amino acids in SAMP1 with strong hydrophobic amino acids isoleucine(Ile) and valine(Val), drastically reduced. Both of the antibacterial activity and the hemolytic toxicity of the peptide SAMP1-A8 increased, in which less strong hydrophobic amino acid tryptophan(Trp) replaced the hydrophobic amino acids in SAMP1.
抗菌肽也称宿主防御肽, 在植物、 昆虫、 两栖动物和哺乳动物等大多数生物体的先天宿主防御机制中起着重要作用, 具有抗菌、 抗病毒及抗肿瘤等作用[1]. 近年来, 抗菌肽被广泛用于生物材料的构建、 生物成像及药物分子的设计等方面[2, 3]. 作为一类抗菌剂, 抗菌肽相对较短(小于50个氨基酸残基), 带正电荷, 具有两亲性(含疏水性和亲水性结构域). 与传统的抗生素相比, 抗菌肽具有抗菌谱广、 热稳定性好及抗菌机理独特等优点[4]. 尽管抗菌肽对各种临床菌株具有很高的抗菌活性, 并且不易产生耐药性, 但其稳定性差、 溶血毒副作用强等缺点限制了其作为临床药物的应用[5, 6, 7]. 因此, 为获得高效低毒的抗菌肽, 研究抗菌肽构效关系, 进而对其结构进行优化十分必要.
影响抗菌肽抑菌活性的2个关键参数是电荷和疏水性[8], 这2个参数在抗菌肽与细菌和哺乳动物细胞膜发生相互作用过程中发挥重要作用, 其中疏水性尤为重要, 会影响抗菌肽进入脂质双分子层的程度[9, 10]. Yin等[11]研究表明, 净电荷量和中度疏水性对抗菌肽的抗菌活性非常重要, 而高疏水性和两亲性与高度溶血性相关. 据文献[12, 13]报道, 精氨酸(Arg)对膜具有很强的亲和力, 序列中富含Arg的肽更容易与细胞膜发生作用. Wimley等[14]研究表明, 带电和具有疏水残基的不完全分离肽在破坏细菌膜方面具有增加的效力.
抗菌肽SAMP1(VRLLKKKI)是本研究组最新发现的一种具有广谱抗菌活性的多肽, 长度为8个氨基酸残基, 净电荷为+4. 本文用Arg替换SAMP1序列中的赖氨酸(Lys)作为带正电荷的氨基酸残基, 并测定了替换前后抗菌活性和溶血性的改变; 同时改变其两亲性和电荷分布, 以验证抗菌肽的活性是否受到序列重排的影响. 采用异亮氨酸(Ile)、 缬氨酸(Val)、 亮氨酸(Leu)和色氨酸(Trp)4种疏水程度不同的氨基酸替换SAMP1序列中的疏水性氨基酸残基, 研究了疏水性改变对抗菌肽作用的影响. 同时, 通过氨基酸残基替换、 序列重排等手段, 设计并合成了8条新的类似肽. 采用圆二色光谱(CD)测定了SAMP1及其类似肽的二级结构, 并测试这些多肽对革兰氏阳性细菌、 革兰氏阴性细菌和致病真菌的抗菌活性以及对人血红细胞的细胞毒性, 以研究这些多肽的结构-活性关系.
实验所用菌株为本实验室保存. 革兰氏阴性细菌选用大肠杆菌(Escherichia coli)和铜绿假单孢菌(Pseudomonas aeruginosa)为代表; 革兰氏阳性细菌选用枯草芽孢杆菌(Bacillus subtilis)、 单增李斯特菌(Listeria monocytogenes)和金黄色葡萄球菌(Staphylococcus aureus)为代表; 致病真菌选用热带念珠菌(Candida tropicalis)为代表. 哌啶、 N, N-二异丙基碳二亚胺(DIC)、 二氯甲烷(DCM)、 N, N-二甲基甲酰胺(DMF)、 三氟乙酸(TFA)、 三异丙基硅烷(TIPS)、 茴香硫醚(Thioanisole)和1, 3-间苯二甲醚(1, 3-Dimethoxybenzene)均购于阿拉丁试剂有限公司; 多肽合成所需的Rink amide-AM树脂、 Fmoc保护氨基酸和N-羟基苯并三唑(HOBt)购于上海吉尔生化公司; 噻唑蓝(MTT)购于美国Sigma公司; 二甲基亚砜(DMSO)购于天津科密欧公司; 2, 2, 2-三氟乙醇(TFE)和十二烷基硫酸钠(SDS)购于上海生物工程有限公司.
FD-1A-50型冷冻干燥机(北京博医康公司); GF-M3000型酶标仪(山东高密彩虹分析仪器有限公司); VS-1300L-U型超净工作台(苏州安泰净化设备有限公司); 安捷伦1260型高效液相色谱仪(HPLC, 美国安捷伦公司); MOS-450多功能圆二色光谱仪(CD, 法国Bio-Logic公司); LCQ Fleet型离子阱液相色谱-质谱联用仪(美国Thermo Fisher Scientific公司).
1.2.1 多肽的合成 参照文献[15]方法, 采用Fmoc固相合成法合成所需多肽, 由羧基端至氨基端合成. 采用反相高效液相色谱(RP-HPLC)梯度洗脱法纯化多肽, 经冷冻干燥获得纯品, 所有多肽纯度大于90%, 于-20 ℃环境下保存. 采用离子肼质谱确定多肽分子量.
1.2.2 生物信息学软件预测理化性质 利用http://www. expasy. ch/tools/网站ProtParam工具分析抗菌肽SAMP1及其类似肽的分子量、 等电点(pI)、 半衰期、 总平均疏水性和脂肪系数等理化性质.
1.2.3 圆二色光谱测定 分别用体积分数为50%的TFE溶液、 溶液SDS(20 mmol/L)和PBS溶液(20 mmol/L)配制终浓度为0.25 mg/mL的抗菌肽SAMP1及其类似肽溶液. CD谱测试条件: 测试温度25 ℃, 石英样品池路径1.0 mm, 响应时间2 s, 步长1 nm, 在波长190240 nm范围内进行扫描, 扫描速度100 nm/min, 每个样品重复测量3次, 取平均值计算平均残基摩尔椭圆率(θ ):
式中: θ obs(mdeg)为CD谱样品检测值减去空白对照值得到的样品椭偏率; l(mm)为石英比色皿的路径长度; c(mmol/L)为肽的浓度; n为多肽所含氨基酸残基个数.
1.2.4 最小抑菌浓度(MIC)的测定 参照文献[16]报道的微量稀释法测定多肽对供试菌的最小抑菌浓度(MIC). 将测试样品在37 ℃培养箱内孵育16 h, 观察孔内溶液浑浊情况. 以96孔板同排中肉眼所见澄清孔所对应的最小浓度为该多肽的MIC值. 真菌MIC测定使用SDA培养基于28 ℃培养过夜.
1.2.5 溶血活性的测定 参考文献[17]方法并加以改进测定溶血活性. 分别称取各多肽, 用生理盐水溶解, 浓度为1 mg/mL, 依次2倍稀释, 加入到96孔板中, 每孔100 μ L, 每个样品平行3次(终浓度分别为500, 250, 125, 64, 32, 16, 8, 4, 2和1 μ g/mL). 将来自健康志愿者的红细胞(RBC)用生理盐水配制成体积分数为2%的红细胞悬液. 取100 μ L红细胞悬液依次加入到上述96孔板各孔中. 用含红细胞体积分数为1%的生理盐水和蜂毒肽(Melittin)分别作为阴性对照和阳性对照. 于37 ℃下孵育1 h, 待红细胞沉淀, 吸取100 μ L上层清液加入到另一个干净的96孔板中, 采用酶标仪在570 nm波长下测定吸光度, 采用公式[(样品OD570-阴性对照OD570)/(阳性对照OD570-阴性对照OD570)]× 100%计算溶血率(%)平均值.
设计合成了一系列类似肽, 重点解析了氨基酸残基组成、 位置以及疏水性改变对抗菌活性和溶血毒性的影响. 选择本实验室筛选得到的具有广谱抗菌活性的抗菌肽SAMP1为模板, 通过序列重排得到类似肽SAMP1-A1, SAMP1-A2和SAMP1-A3, 其序列中氨基酸残基组成和个数不变, 进一步研究了氨基酸残基位置改变对抗菌肽的影响. 利用精氨酸(Arg)取代SAMP1序列中的赖氨酸(Lys)得到类似肽SAMP1-A4, 研究了带正电荷氨基酸残基改变造成的影响. 异亮氨酸(Ile)、 缬氨酸(Val)和亮氨酸(Leu)是母肽SAMP1序列中带有的3种疏水性氨基酸残基. 据文献[18]报道, 在细菌和动物体内发现的抗菌肽序列中, 疏水性氨基酸残基Ile, Val和Leu出现的频率较高. 鉴于三者总体疏水性均较强, 故选择色氨酸(Trp)作为较弱疏水性氨基酸残基的代表. 基于此, 选用疏水强度不同的Ile, Val, Leu和Trp替换SAMP1序列中的疏水性氨基酸残基, 得到类似肽SAMP1-A5, SAMP1-A6, SAMP1-A7和SAMP1-A8, 进一步研究了疏水性改变对抗菌肽效用的影响. 实验所用多肽序列见表1.
| Table 1 Amino acid sequences, molecular mass and HPLC retention time of SAMP1 and its analog peptides |
Mojsoska等[19]将HPLC峰保留时间(tR)与多肽疏水性相关联, 发现在相同的液相条件下, 疏水性强的多肽具有更高的保留时间. 由表1可知, 类似肽SAMP1-A5的tR值最大, 表明其疏水性最强; 而类似肽SAMP1-A8的tR值最小, 表明其疏水性最弱. 经离子肼质谱分析确定所合成多肽的分子量正确.
ProtParam是一种生物信息学蛋白预测分析软件, 可以根据输入的蛋白质序列预测各种物理和化学参数. 抗菌肽SAMP1及其类似肽的理化性质预测结果如表2所示.
| Table 2 Physicochemical properties of SAMP1 and its analog peptides |
总平均疏水性(GRAVY)是指蛋白序列中氨基酸残基亲疏水值相加除以氨基酸残基总数得到的比值, 负值越大代表亲水性越强, 正值越大代表疏水性越强. 从预测结果可知, 类似肽SAMP1-A4, SAMP1-A6和SAMP1-A8均为亲水性肽, SAMP1-A8亲水性最强; 其余类似肽均为疏水性肽, 且SAMP1-A5疏水性最强. 通常, 稳定系数< 40表示蛋白稳定性较高; 稳定系数> 40则表示蛋白不稳定. 类似肽SAMP1-A4和SAMP1-A8经预测均为不稳定多肽. 半衰期是指多肽在细胞中降解一半时所需要的时间. 经过预测SAMP1-A2, SAMP1-A3, SAMP1-A6和SAMP1-A8半衰期较短. 将N端氨基酸残基Val替换为Arg后, 通过预测发现类似肽SAMP1-A2和SAMP1-A3的半衰期显著降低, 证明N端氨基酸残基种类对多肽半衰期影响很大.
CD光谱通常用于确定和研究蛋白的二级结构. 分别以PBS溶液为水相环境, SDS溶液为负电荷环境, 体积分数为50%的TFE溶液为疏水环境, 对SAMP1及其类似肽的二级结构进行了CD谱测定, 结果见图1.
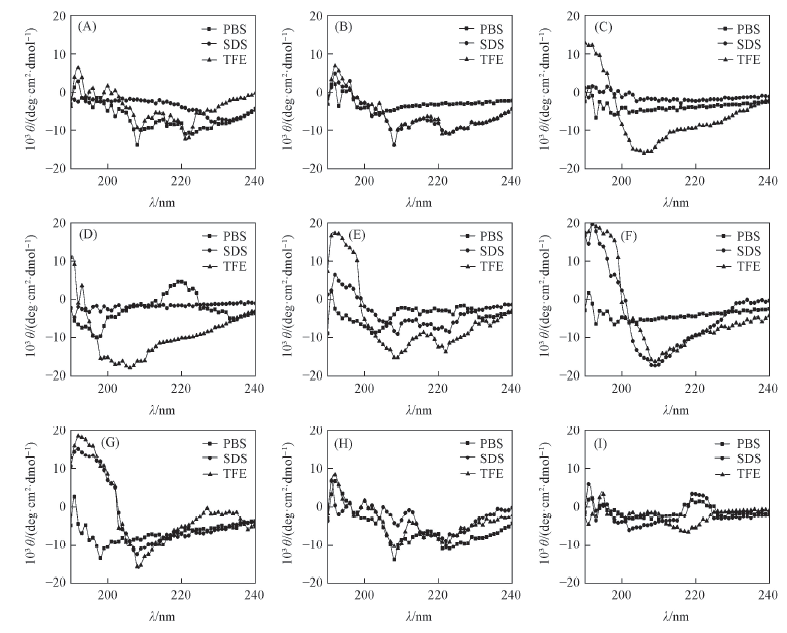 | Fig.1 CD spectra of SAMP1 and its analog peptides in PBS, SDS and TFE solutions (A) SAMP1; (B) SAMP1-A1; (C) SAMP1-A2; (D)SAMP1-A3; (E) SAMP1-A4; (F) SAMP1-A5; (G) SAMP1-A6; (H) SAMP1-A7; (I) SAMP1-A8. |
CD谱图中192 nm附近形成正峰, 并在208和222 nm附近出现2个负特征峰, 这是典型的α 螺旋谱图. 由图1可知, 当大部分类似肽处于体积分数为50%的TFE溶液中时, 192 nm处的正峰和208, 222 nm处的2个负特征峰强度均得以增强, 证明α 螺旋含量增加. 利用CDPro软件中的SELCON3, CDSSTR和CONTIN程序分别计算了各二级结构要素含量, 结果列于表3. 可见, SAMP1及大部分类似肽二级结构以α 螺旋和无规则卷曲为主. 在50%(体积分数)TFE溶液中, α 螺旋比例增加. 大部分类似肽α 螺旋含量相近, 表明多肽穿膜过程以α 螺旋结构为主, 但其抗菌活性与α 螺旋含量并不直接相关.
| Table 3 Secondary structure content of SAMP1 and its analog peptides |
SAMP1及其类似肽对热带念珠菌、 革兰氏阳性细菌和革兰氏阴性细菌的抗菌MIC测试结果列于表4. 可见, 抗菌肽SAMP1及其类似肽对热带念珠菌抑菌活性变化不大. 经过序列重排后, 类似肽SAMP1-A1, SAMP1-A2和SAMP1-A3对枯草芽孢杆菌抗菌活性提高815倍, 但对铜绿假单胞菌抑菌活性降低832倍. 研究表明, 精氨酸残基比赖氨酸残基具有更强的静电结合能力, 替代抗菌肽中的精氨酸残基可显著减少细菌杀伤[20, 21, 22]. 本文用Arg替换Lys得到的类似肽SAMP1-A4与SAMP1相比具有更高的广谱抗菌活性, 对革兰氏阳性枯草芽孢杆菌和金黄色葡萄球菌的抑菌活性提高3060倍, 这与文献[20, 21, 22]结果一致. 疏水性是影响抗菌肽生物活性的关键参数. Gopal等[23]研究表明, 序列总体疏水性超越一定范围后, 可导致抗菌肽的抗菌活性降低. 本文用Ile和Val分别作为序列中的疏水性氨基酸残基, 得到的类似肽SAMP1-A5和SAMP1-A7对革兰氏阳性细菌和革兰氏阴性细菌的抑菌活性均大幅度降低(MIC> 250 μ g/mL), 但对真菌的抑菌活性影响不大; 用Leu和Trp作为疏水性氨基酸残基得到的类似肽SAMP1-A6和SAMP1-A8均表现出较强的广谱抑菌活性. 上述结果表明, 疏水性并不是越强越好, 存在一定的最佳范围, 超过阈值会导致抗菌活性急剧降低. 这种现象可以通过肽自身缔合来解释, 肽自身缔合会阻止其在原核细胞中穿过细胞壁, 而这种肽自缔合不影响真核细胞膜的进入.
| Table 4 Antimicrobial activity of SAMP1 and its analog peptides |
为了评估肽对哺乳动物细胞的细胞毒性, 测定了不同浓度肽对人血红细胞的溶血百分比. 表5结果显示, 经过序列重排后正电荷氨基酸残基聚集在C端的类似肽SAMP1-A1和正电荷氨基酸残基均匀分布的类似肽SAMP1-A2的溶血毒性升高; 而正电荷氨基酸残基聚集在N端的类似肽SAMP1-A3的溶血毒性降低. 用Arg替换Lys得到的类似肽SAMP1-A4与抗菌肽SAMP1相比溶血毒性降低. 用Val作为疏水性氨基酸残基的类似肽SAMP1-A7的溶血性降低; 而用Trp作为疏水性氨基酸残基的序列SAMP1-A8则显示出溶血毒性增加, 表明序列中加入色氨酸残基可能会引起人红细胞溶血.
| Table 5 Hemolysis ratio of SAMP1 and its analog peptides |
基于抗菌肽SAMP1合成了8种类似肽, 旨在探究氨基酸残基位置、 种类和疏水性变化对多肽功能的影响. 结果表明, 正电荷氨基酸残基的位置对抗菌肽抗菌活性的影响不大, 但对溶血性有影响. 用Arg替换Lys能提高抗菌肽对革兰氏阳性细菌的抑菌活性, 同时降低溶血性. 疏水性较强可能导致抗菌肽抑菌活性降低. 用Leu和Trp作为序列中的疏水性氨基酸残基, 可使抗菌肽抗菌活性升高, 但是Trp的取代使抗菌肽溶血毒性增加. 经结构改造, 得到的类似肽SAMP1-A3和SAMP1-A4具有广谱的高抑菌活性, 同时溶血毒性低, 具有进一步研究价值.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|